"Arbuscular mycorrhization (AM)" is a root endosymbiosis between over 80% of plants and Glomeromycota fungi. AM development is accompanied by an exchange of signalling molecules between plant and mycorrhizal fungi. The first signals proposed are strigolactones released from the plant roots and perceived by mycorrihizal fungi in the rhizosphere. Mycorrhizal fungi release signalling molecules, called Myc factors which trigger root symbiotic response, such as calcium spiking and marker gene expression. Once the dialogue has been made between the fungus and the plant, roots and hyphae will proliferate and branch in the rhizosphere. When a hyphal tip touches the surface of a root, it receives signalling from host plant and will swell, flatten on the cell wall of an epidermal cell and branch repeatedly to develop what is known as a hyphopodium. Meantime the epidermal cells contacted with the fungus start to reorganize their cytoplasm to produce an AM-specific structure, called the PPA (prepenetration apparatus) which is required for successful fungal penetration. Once the fungus penetrates into host root and reaches into the inner cortical cells, where the fungus will form a highly branched structure called an arbuscule, the site of nutrient exchange between plant and fungus.
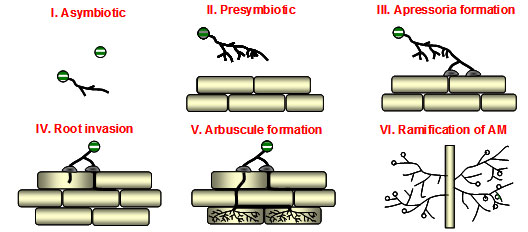
Stages of infection of mycorrhizal fungi
We have identified RAM, a gene which is required for early fungal (mycorrhizas and pathogens) hyphopodia/appressoria formation on the root surface and arbuscule development in cortical cells. RAM appears to have a direct signalling function at least at the root surface, since exogenous addition of lipids is sufficient to promote appressoria formation. The phenotype of ram showed that the RAM gene functions both at the epidermis and the cortical cells, interestingly RAM was highly induced in arbuscules, the late stage of this symbiosis, suggesting that fatty acids may also serve nutritional function in the late stage of this symbiosis. It would be very important to design experiments to prove this hypothesis in future. Another important issue worth pursuing is to identification which kinds of lipid molecules are most active for fungal hyphopodia formation and how plant lipid metabolism is regulated during mycorrhizal fungi infection. In the short term, it will be interesting to dissect the RAM promoter and define the regions responsible for mycorrhizal specific expression and then use this cis-element to identify transcription factors that are necessary for its activation using yeast one hybrid approach. We are also continuing to explore the role of these lipids in the early and late stages of the AM symbiosis through reverse genetic approaches combined with cell biology, bioinformatics and metabolome data.
We have known that the infection process is highly regulated by the host plant. In the long term, we would like to focus on understanding how this symbiotic progress is tightly regulated by host plant. We have recently isolated five aam (Abolished Arbuscular Mycorrhizal formation) Medicago mutants through forward genetic screening for AM symbiosis defective phenotypes, which showed wild-type responses to Sinorhizobium meliloti infection. Cloning and characterization of these AAM genes will deliver knowledge about how mycorrhizal fungal colonization is regulated by plant.
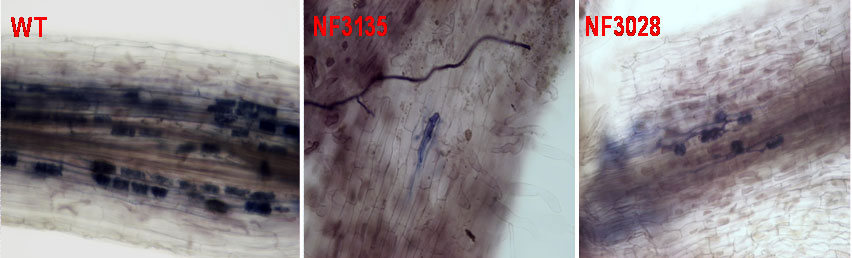
Two abolished arbuscular mycorrhizal formation mutants in Medicago